By Maartje De Vos, Researcher functional ingredients
Giving birth and being born is tough
Birth is a serious challenge for any mammal, and swine are no exception. In fact, because litter size has been increased dramatically in recent decades through genetic selection, birth complications have also increased. Although sows appear fairly calm prior to parturition, metabolically there is tremendous activity the hours preceding farrowing (Kelley, 1978). Farrowing itself also taxes the sow enormously. Consequently, sows, especially with very large litters, can become physically exhausted. When this happens, the birth process is interrupted – a process called dystocia. This is especially harmful in litter-bearing animals: fetuses that are in the process of being born can become suffocated. A long period of suffocation leads to stillbirths; and a shorter period of suffocation harms the vitality of the newborn piglet – it struggles to get up and drink colostrum. And not only the fetuses suffer from dystocia; the sow is harmed as well, including increased incidence of mastitis, metritis, and agalactia (MMA). Ergogenic aids administered early in parturition have resulted in fewer health complications in sows (van Kempen, 2007).
Newborn piglets have another handicap that is often overlooked: their metabolic rate is notably lower than what textbooks claim (about 40% lower; van Kempen, 1993). This is likely because the digestive system is still in standby modus, awaiting the first food to activate it. Moreover, being born wet and in a relatively cold environment increases the chances of hypothermia. Therefore, it is critical for piglet immunity to start eating as soon as possible in order to boost its metabolic activity and thus heat production.
Being born with moderate hypoxia and a very low metabolic rate are two strikes against the newborn pig. In practice, this can often be fatal, unless management is very alert. Interestingly, Otten (1994) used low-intensity microwaves to reheat hypothermic piglets that were doomed to die. Once rewarmed, the piglets regained vitality and were able to nurse on their own. This underscores the fact that hypothermia is a key problem, not the piglet’s limited energy reserves as is often claimed.
Colostrum – life elixir
Colostrum is very rich in nutrients as well as in immunoglobulins, antimicrobial peptides, and growth factors, and it is critical for a healthy start. The importance of drinking adequate colostrum was nicely demonstrated by Shaw (Figure 1; 1989) – when exposed to rotavirus, piglets that did not receive colostrum experienced 80% morbidity and 60% mortality, while piglets that received colostrum experienced zero morbidity and mortality. Colostrum consumption is also key for heat production. Gentz (1971) showed that, when piglets nurse, their heat production normalizes.
Immune challenges for the piglet
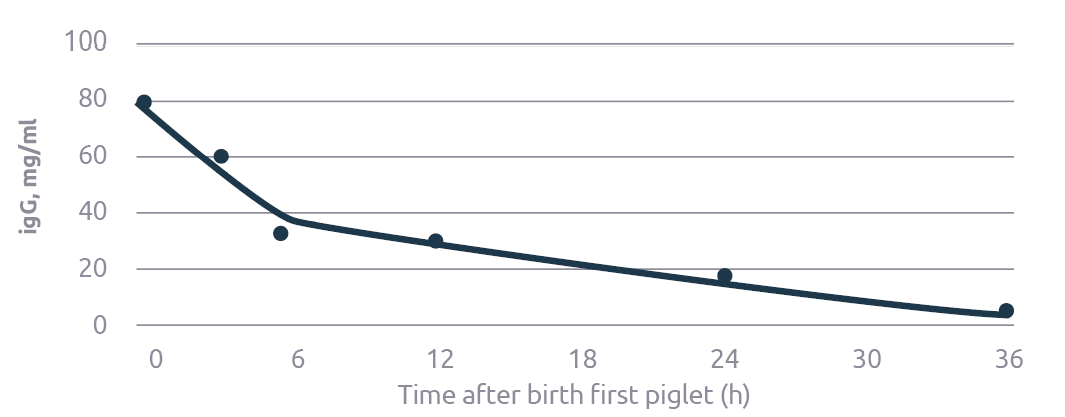
Piglets face an obstacle course immunologically as well. Because the sow’s placenta does not allow for the transfer of immunoglobulins, newborn piglets lack a key component of immune health. Colostrum is supposed to supply the piglet with the necessary immunoglobulins, but colostrum is in limited supply and immunoglobulin levels drop exponentially during the first hours of lactation (Figure 2). Piglets born late within a litter are more prone to dystocia, which harms their ability to fight for colostrum. On top of that, their somewhat older siblings may well have depleted colostrum to the point that the late-born piglets are unable to obtain sufficient colostrum, even when they were born fit or when they received help.
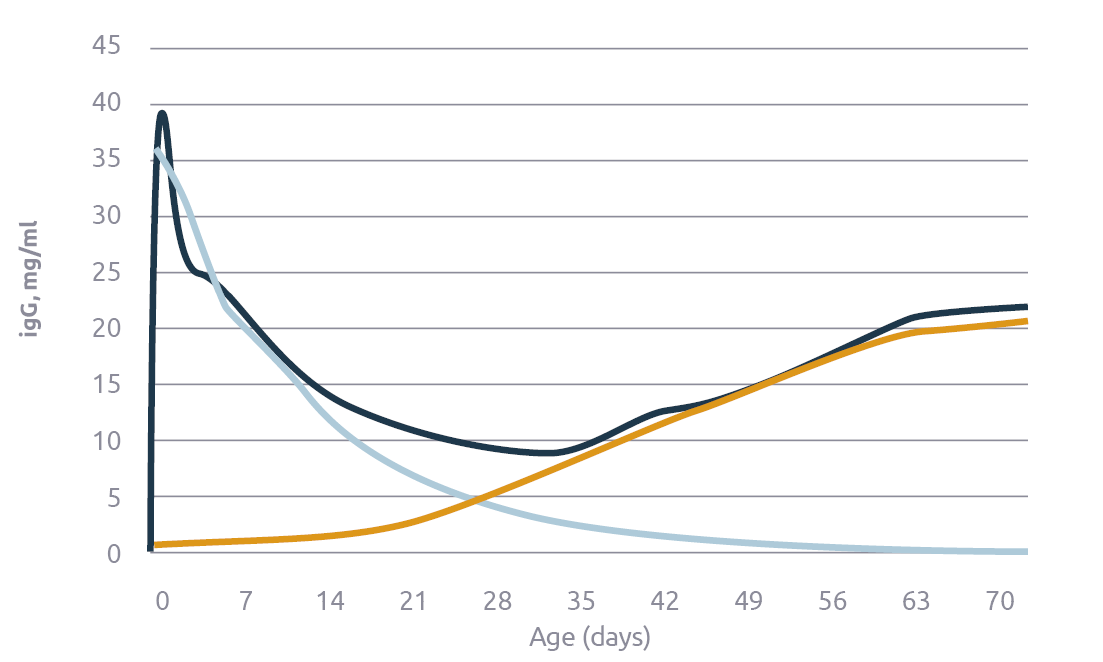
Even when piglets get a healthy dose of immunoglobulins with colostrum, these maternal immunoglobulins are typically catabolized at 3 to 4 weeks of age – the age at which we typically wean the piglets. However, the piglet’s own immune system is slow to mature, and immunoglobulin production has barely started at weaning (Polo, 2012). So, we usually wean piglets when they experience an immunoglobulin dip (Figure 3).
Abrupt weaning of piglets results in stress and a period of food-withholding, which perturbs intestinal health. For decades, this was managed with antibiotics, copper, and zinc oxide, which took the pressure off the piglet’s immature immune system. These additives are now deemed unsustainable and are consequently banned (for example, in the EU), and natural products with antimicrobial properties have been developed as alternatives. These include organic acids, essential oils, and medium-chain fatty acids (MCFA), as well as high-quality diets aimed at maximizing feed intake. The industry is to be congratulated for these efforts, as antibiotic usage has decreased 10-fold in countries that have been working on this topic in recent years (SDa autoriteit, 2023)! Nevertheless, despite great strides, health problems including diarrhea and Streptococcus suis infections remain important challenges in nursery piglets.
Improving piglet immunity starts by supporting their mother optimally
Strategies for improving the survival of piglets around the birth process ideally focus on the source of the problems: and this is foremost the sow, not the piglets. The sow should be prepared for farrowing by maximizing glycogen reserves and endurance. But we need to keep in mind that the energy demand for fetal growth increases exponentially in the days before farrowing – therefore, moving sows shortly before farrowing can stress them out, resulting in glycogen depletion with insufficient time to recover. Thus, a rather high feed gift is required (Cools, 2014) to allow the sow to support the gain of her litter, the reproductive organs (in preparation for farrowing), the mammary gland including colostrum, and, on top of that, glycogen restoration. Stress compounds all of these things. Boosting endurance is a tough challenge in practice. Group housing of gestating sows may well be a small step in the right direction.
Immune support for the piglet
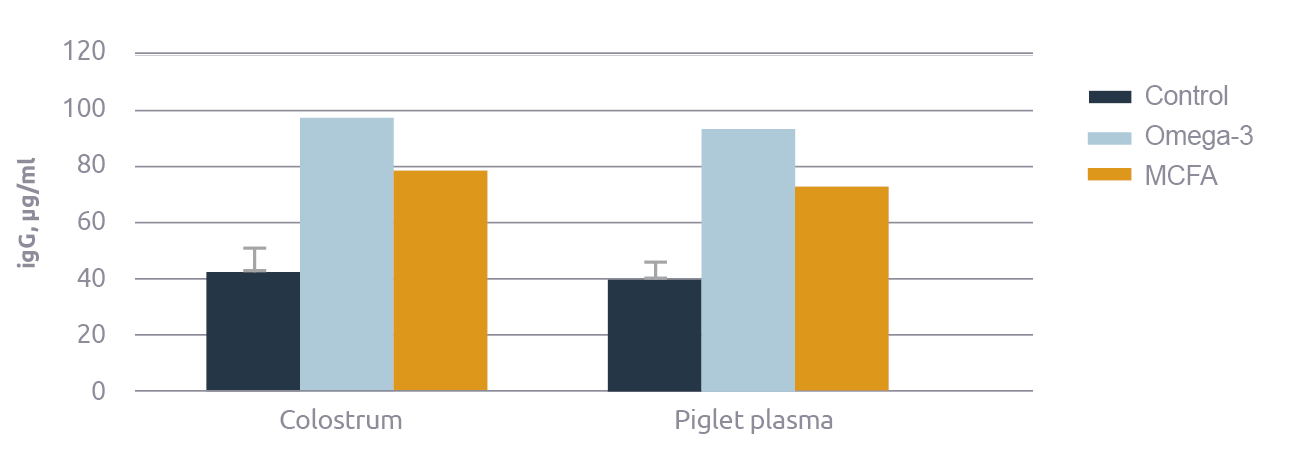
We can help the sow better prepare for farrowing immunologically as well. For example, omega-3 fatty acids are anti-inflammatory and, through that mechanism, they can strongly boost the immunoglobulin content of colostrum (Figure 4; Chen, 2019). Being anti-inflammatory, however, has a downside: inflammation is a critical response for combatting intracellular pathogens like viruses (Klasing, 2007). So, playing with the ratio of omega-3/omega-6 fatty acids is yin-yang: the outcome is very situation-dependent. With exposure to intracellular pathogens like PRRSv, pro-inflammatory omega-6 fatty acids may well be preferred; but with extra-cellular pathogens like E. coli, anti-inflammatory omega-3 fatty acids are preferred (Husson, 2016). In line with this, Walter et al. (2019) demonstrated that the pro-inflammatory arachidonic acid (omega-6) improves health in LPS-challenged piglets.
Medium-chain fatty acids improve piglet immunity
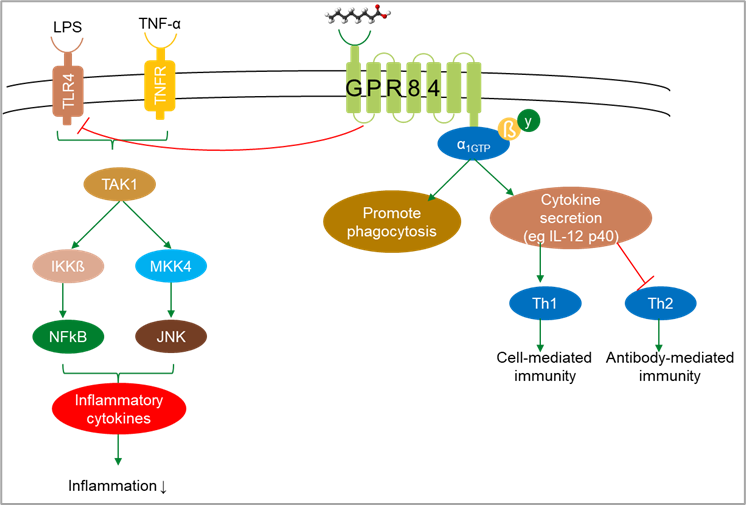
A fairly recent discovery is that MCFA can also interact with piglet immunity. They do so via a completely different mechanism than omega-3 fatty acids; they can stimulate G-protein-coupled receptors (GPR), especially GPR84, present on immune cells (Roopashree, 2021). This activation is most pronounced with decanoic acid (C10; Wang, 2006). Activation of GPR84 by MCFA inhibits responses to LPS and to TNF-α and promotes phagocytosis, cell-mediated immunity, and antibody-mediated immunity (Figure 5).
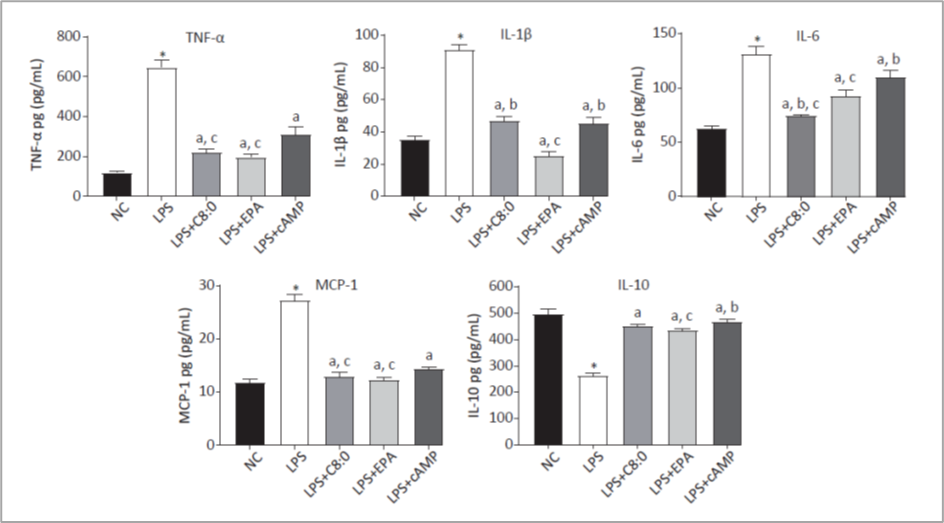
Indeed, despite the differences in mechanisms, MCFA and eicosapentaenoic acid (EPA) yielded very similar effects in vivo and in macrophages stimulated by LPS (Figure 6); they both suppressed pro-inflammatory cytokines (TNF-α, IL-1ß, IL-6, and MCP-1), and they both stimulated anti-inflammatory cytokines (IL-10; Zhang, 2022). This boost of immune health results in higher levels of immunoglobulin A (Kono, 2004) and immunoglobulin G in both the sow’s blood (Figure 4; +59%) and colostrum (+80%; Chen, 2019). MCFA also increased the health of polymorphonuclear leucocytes – a primary mediator of innate immunity – in nursery piglets.
Through these GPR84 receptors, MCFA can also boost mammary gland development (Meng, 2017) and thus colostrum production. Litter weight gain the first 24 hours after farrowing is indeed higher in sows that are fed MCFA, indicating that colostrum output is improved by MCFA (internal data).
After weaning, MCFA can do double duty by both directly inhibiting pathogens like E. coli as well as indirectly affecting pathogens through boosting immune health as explained above. Indeed, under controlled infection experiments, MCFA decreased the susceptibility of piglets to co-infection by PRRSv and S. suis applied nasally (Figure 8). Under field conditions, MCFA decreased the severity of a PRRS outbreak (decreasing mortality from 17.8 to 5.9%; PMI) and the complications arising from endemic S. suis.
MCFA: a versatile tool for boosting health and piglet immunity
Thus, MCFA are advantageous not only for combatting pathogens in the early part of the intestinal tract through their detergent properties, but also for boosting immune health. When fed to gestating sows, MCFA can boost both colostrum Ig content as well as total colostrum output. This means that piglets born later in the litter have a better chance of consuming sufficient colostrum in order to reach their required immunoglobulin limit, and thus enjoy a healthy start of their life. When fed to nursery piglets, MCFA increase piglet immunity and the piglet’s ability to fight off both bacterial pathogens like S. suis as well as viral pathogens like PRRSv. In sum, the versatility of MCFA for boosting animal health keeps expanding.